The properties of a polymer change significantly depending on the structure of the polymer chain, particularly, with branched structures, depending on the number of branches and the length of the branch.* However, the long-chain branch (LCB) structure of polyethylene was unclear, due particularly to the complex polymer structure and the limitations of its analysis methods.
In their study “Direct Observation of Long-Chain Branches in a Low-Density Polyethylene” Ken-ichi Shinohara, Masahiro Yanagisawa and Yuu Makida measured the chain length of LCBs and the distance between branch points of LDPE by atomic force microscopy.*
Figure 1 from “Direct Observation of Long-Chain Branches in a Low-Density Polyethylene “ by K. Shinohara et al.: Direct measurement of LCB in a tubular LDPE (F200-0 fractionated). (A) AFM image of a single molecule of LDPE on mica in DMTS at 25 °C. X: 279 nm, Y: 209 nm, Z: 18 nm. (B) Length of each chain of LDPE. (C) A wire model of self-shrinking structure of polymer chain of LDPE. Main chain: red wire. LCB: black wire. The model was created to be one tenth of the length of the extended chain based on AFM observation (B), MD simulation (Fig. S1), and the molecular weight determined by SEC-MALLS-Visc experiments (see Fig. 2).
*Ken-ichi Shinohara, Masahiro Yanagisawa, Yuu Makida Direct Observation of Long-Chain Branches in a Low-Density Polyethylene Nature Scientific Reportsvolume 9, Article number: 9791 (2019) doi: https://doi.org/10.1038/s41598-019-46035-9
Open Access: The article «Direct Observation of Long-Chain Branches in a Low-Density Polyethylene» by Ken-ichi Shinohara, Masahiro Yanagisawa and Yuu Makida is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Mucosal immunoglobulins comprise mainly secretory IgA antibodies (SIgAs), which are the major contributor to pathogen-specific immune responses in mucosal tissues. SIgAs exist as mainly dimers and tetramers and play critical roles in mucosal immune responses against influenza.*
Detailed characterization of these anti-viral SIgA is important for better understanding of the mechanisms underlying anti-viral immunity.* In their article “IgA tetramerization improves target breadth but not peak potency of functionality of anti-influenza virus broadly neutralizing antibody” Saito S, Sano K, Suzuki T, Ainai A, Taga Y, Ueno T, et al. (2019) describe a means of generating a recombinant tetrameric monoclonal SIgA to enable exhaustive characterization of tetrameric SIgAs. The tetrameric monoclonal SIgA possessing variable regions of anti-influenza viruses broadly neutralizing antibody show that tetramerization of SIgA improves target breadth, but not the peak potency, of their anti-viral functions.* These results broaden the knowledge about the fundamental role of SIgA tetramerization in anti-viral humoral response at the human respiratory mucosa.*
The high speed atomic force microscopy ( HS-AFM ) experiments mentioned in the article were performed using a NanoWorld Ultra-Short Cantilever USC-F1.2-k0.15.
Fig 1. Production of recombinant tetrameric monoclonal SIgAs from ” IgA tetramerization improves target breadth but not peak potency of functionality of anti-influenza virus broadly neutralizing antibody ” by Saito S, Sano K, Suzuki T, Ainai A, Taga Y, Ueno T, et al. (2019) :
(A) Recombinant monoclonal IgA antibodies purified from the culture supernatant of cells co-transfected with A1+L (left upper), A1+L+J (left lower), A1+L+J+SC (right upper), or A2m2+L+J+SC (right lower), were subjected to size exclusion chromatography (SEC) analysis. A chromatogram showing absorbance at 280 nm revealed three major peaks: peak A (retention volume around 10.4 ml), peak B (retention volume around 9.3 ml), and peak C (retention volume around 8.4 ml). Data are representative of three independent experiments. (B) SDS-PAGE and BN-PAGE analysis of IgG and IgA1/IgA2m2 in each peak fraction (peak A, B, and C) purified from cells co-expressing SC (A1+L+J+SC or A2m2+L+J+SC). (C, D, E) High-mass MALDI-TOF MS analysis of the each peak fraction containing recombinant IgA1 purified from the culture supernatant of cells transfected with A1, L, J, and SC. (C) One main peak (arrow) corresponding to monomer (Mo) was detected in the peak A fraction. (D) Two main peaks (arrows) corresponding to a dimer (Di) and a di-cation dimer (Di2+) were detected in the peak B fraction. (E) Three main peaks (arrows) corresponding to a tetramer (Te), trimer (Tr), and di-cation tetramer (Te2+) were detected in the peak C fraction. (F, G) High-mass MALDI-TOF MS analysis of the each peak fraction of recombinant IgA2m2 purified from the culture supernatant from cells transfected with A2m2, L, J, and SC. (F) One main peak (arrow) corresponding to a monomer (Mo) was detected in the peak A fraction. (G) Three main peaks (arrows) corresponding to a tetramer (Te), a trimer (Tr), and a di-cation tetramer (Te2+) were detected in the peak C fraction. (H) Quantification of the amount of each subunit within the peak B or C fraction of recombinant SIgA1 or SIgA2m2 antibodies purified from the culture supernatant of cells transfected with A1/L/J,/SC or A2m2/L/J/SC using LC-MS with stable isotope-labeled standard peptides. The abundance of each subunit to that of J chain is expressed as a ratio. Data are expressed as box-and-whisker plot with minimum, maximum, median, upper and lower quartiles (n = 6–7). (I) HS-AFM image of peak C derived from a recombinant SIgA1 (A1Te) or SIgA2m2 (A2m2Te) antibody purified from the culture supernatant of cells transfected with A1/L/J/SC or A2m2/L/J/SC. Scale bar, 20 nm.
Open Access: The article « IgA tetramerization improves target breadth but not peak potency of functionality of anti-influenza virus broadly neutralizing antibody » by Saito S, Sano K, Suzuki T, Ainai A, Taga Y, Ueno T, et al. (2019) is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
In their recent publication “High-speed AFM height spectroscopy reveals μs-dynamics of unlabeled biomolecules” in Nature Communications George R. Heath and Simon Sheuring develop and apply HS-AFM height spectroscopy (HS-AFM-HS, a technique inspired by fluorescence spectroscopy), a technique whereby the AFM tip is held at a fixed x–y position and the height fluctuations under the tip in z-direction with Angstrom spatial and 10µs temporal resolution are monitored.
They demonstrate “how this technique can be used to simultaneously measure surface concentrations, diffusion rates and oligomer sizes of highly mobile annexin-V molecules during membrane-binding and self-assembly at model membranes and derive its kinetic and energetic terms. Additionally, HS-AFM-HS at specific positions in the annexin lattice where the freedom of movement is restricted to rotation allowed determination of the interaction free energies of protein-protein contacts.”* The applicability of this technique is wide and is discussed at the end of the publication.
NanoWorld Ultra-Short Cantilevers (USC) for Fast-/High-Speed AFM ( USC-F1.2-k0.15 ) were used.
Congratulations to the authors to this publication which pushes the speed limits of AFM even further!
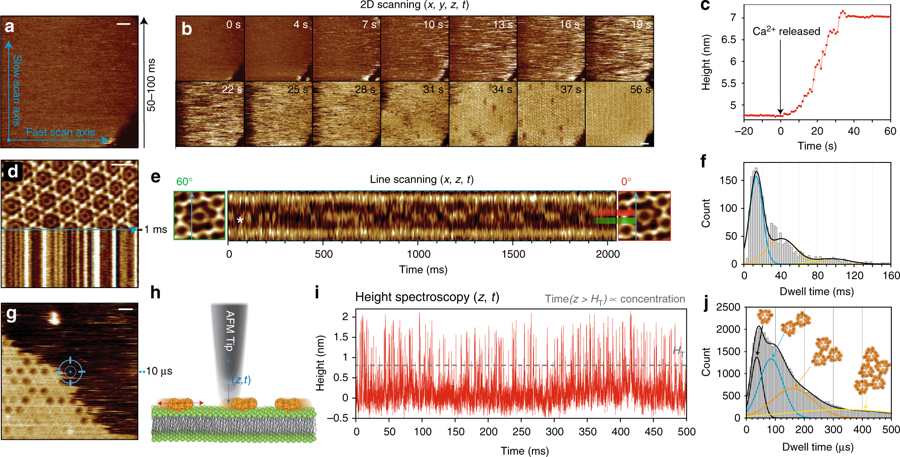
Figure 1 from “High-speed AFM height spectroscopy reveals μs-dynamics of unlabeled biomolecules”: Increasing the temporal resolution of HS-AFM by reducing the dimensionality of data acquisition. a HS-AFM image of a DOPC/DOPS (8:2) membrane in the presence of annexin-V and NP-EGTA-caged Ca2+. Blue arrows illustrate the slow- (vertical) and the fast-scan axis (horizontal). Images can be captured at up to 10–20 frames s−1. b HS-AFM movie frames of A5 membrane-binding, self-assembly and formation of p6 2D-crystals upon UV-illumination induced Ca2+-release. c Average height/time trace of the membrane area in b. d Averaged HS-AFM image of an A5 p6-lattice overlaid with the subsequent line scanning kymograph, obtained by scanning repeatedly the central x-direction line as illustrated by the blue arrow with a maximum rate of 1000–2000 lines s−1. e Line scanning kymograph across one protomer of the non-p6 trimer, marked by * in d and e at a rate of 417 lines s−1 (2.4 ms per line). f Histogram of state dwell-times of the molecule in e. g HS-AFM image of an A5 p6-lattice partially covering a DOPC/DOPS (8:2) SLB surface during self-assembly. HS-AFM height spectroscopy (HS-AFM-HS) is performed following halting the x- and y-piezos to capture height information at a fixed position at the center of the image (illustrated by the target). h Schematic showing the principle of HS-AFM-HS. The AFM tip is oscillated in z at a fixed x,y-position, detecting single molecule dynamics such as diffusion under the tip. i Height/time trace obtained by HS-AFM-HS with the tip positioned at the center of image (g). The height/time trace allows determination of the local A5 concentration analyzing the time fraction of the occurrence of height peaks. j Dwell-time analysis of each height peak of diffusing A5 from 60 s height/time data and subsequent fitting of the distribution to multiple Gaussians (possible molecular aggregates corresponding to the fits with distinct dwell-times (τD) are shown above the graph). All scale bars: 20 nm
*George R. Heath & Simon Scheuring High-speed AFM height spectroscopy reveals μs-dynamics of unlabeled biomolecules Nature Communicationsvolume 9, Article number: 4983 (2018)
DOI: https://doi.org/10.1038/s41467-018-07512-3
Open Access The article “High-speed AFM height spectroscopy reveals μ s-dynamics of unlabeled biomolecules” by George R. Heath & Simon Scheuring is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.